Restoring Oak Woodlands in California: Theory and Practice  2001
2001
Phytosphere Research, 1027 Davis Street, Vacaville, CA, 95687-5495
e-mail: phytosphere@phytosphere.com
Copyright 2001 Phytosphere Research - All Rights Reserved
Ecological importance of California oak woodlands
Vegetation types dominated by oak trees cover about 4 million hectares in California (Bolsinger 1988), or roughly 10% of the state’s land area. These extensive oak woodlands serve a number of important ecological functions. Oak woodlands play a critical role in protecting soils from erosion and landsliding, regulating water flow in watersheds, and maintaining water quality in streams and rivers. Oak woodlands also have higher levels of biodiversity than virtually any other terrestrial ecosystem in California. At least 300 terrestrial vertebrate species (Block, Morrison, and Verner 1990), 1,100 native vascular plant species (CalFlora Database 1998), 370 fungal species and an estimated 5,000 arthropod species (Swiecki et al. 1997a) are associated with California oak woodlands.
More than twenty-five oak species, natural hybrids, and varieties are native to California. California oaks also occur as components in desert plant communities and conifer-dominated montane and forest ecosystems. In this chapter, our discussion is limited to the low elevation oak woodlands of valleys and foothills. In these regions, almost all precipitation falls between September and May and seasonal totals vary from about 27 cm in the driest woodlands to 93 cm or more in the more mesic areas. Winter temperatures only occasionally drop to or below freezing. Summer temperatures are moderate near the coast, but most inland areas are subject to intermittent summer temperatures above 38 C. The dominant oaks in these areas are valley oak (Quercus lobata), blue oak (Q. douglasii), interior live oak (Q. wislizeni) and coast live oak (Q. agrifolia). Engelmann oak (Q. engelmannii) replaces blue oak in southern California.
Loss of oak woodlands
California’s oak woodlands have been dramatically reduced in extent over the past 230 years. European and later American settlers greatly reduced woodland cover, most dramatically in the first 50 years after the discovery of gold in California in 1848. Several oak species were extensively harvested for fuelwood and charcoal production, but most California oaks had little or no commercial value as timber. Widespread destruction of oak woodlands to clear land for more profitable uses has been an accepted practice which continues to the present day in many areas. Clearing for intensive agriculture, rangeland "improvement", and urban development have eliminated oak woodlands from much of their former range. Stringers of oaks along creeks and occasional oaks scattered in agricultural fields and on grassy hillsides sometimes provide the only hints of the extent of former woodlands.
Biotic and physical characteristics of former and extant woodlands have also been changed from their presettlement condition. Numerous plant and animal species have been irrevocably lost, while many nonindigenous plant and animal species have become so widespread that their eradication is impossible. Although many native species are still present in these degraded ecosystems, the herbaceous layer has become dominated by nonindigenous annual grasses and forbs. Grazing and clearing have increased soil compaction and erosion. In some areas, hydraulic mining removed entire hills and redeposited their contents downstream. Dams have flooded tens of thousands of hectares of oak woodland and have changed historic water tables and flooding regimes under which riparian oak forests developed. Groundwater pumping and accelerated streambed incision have also lowered water tables in valleys.
Although all oak woodland types have been affected by the changes brought about by European settlement, woodlands dominated by blue, valley, and Engelmann oak have been the most adversely affected. For all of these species, losses due to clearing for agriculture, urbanization, and fuelwood have been compounded by regeneration failure within existing stands. Sapling populations in many oak woodlands are insufficient to offset mortality and maintain current stand densities (Bolsinger 1988, Swiecki et al. 1997c). Without natural regeneration, woodlands have gradually thinned to open oak savannas, which in turn are converted to grasslands dominated by nonindigenous annuals. Studies of the age structure of various oak stands indicate that most existing woodlands are composed primarily of second growth that established between the 1850s and the first decades of the twentieth century (e.g., Mensing 1992). Widespread suppression of oak regeneration, especially in blue oak woodlands, is a relatively recent phenomenon that is largely restricted to lands that have been used for livestock range for many years (Swiecki and Bernhardt 1998).
The need for oak woodland restoration is a consequence of past and current management practices that have degraded or destroyed these ecosystems. Thus, restoration and management of oak woodlands are inseparably linked. For either oak woodland restoration or sustainable management of existing woodlands, we must answer the following questions:
- What do you have? What is the condition of the existing ecosystem and how did it come to its current state?
- What do you want? What outcomes for the ecosystem are both attainable and ecologically appropriate?
- How do you get what you want? What inputs and techniques can be applied to achieve restoration / management goals?
- Are you getting what you want? How do we assess the success of restoration / management efforts?
In this paper, we will consider the theoretical basis for addressing these four questions. We will also discuss practical applications of the principles that follow from these theoretical considerations.
What do you have? - Past and present vegetation and management
Restoration logically begins with an understanding of the condition of the resource and the management history that has brought it to its current state.
1. What is the current composition of the vegetation? Current vegetation is the starting material of a restoration project and may include desirable species to be favored as well as undesirable species to be suppressed or eradicated.
2. What was the nature of the former woodlands? Historical photos, accounts, and herbarium specimens can be used to gain insight into the former character of the woodlands, but it is impossible to definitively reconstruct the presettlement oak woodland vegetation. Nonindigenous plants introduced by the earliest European explorers and early Spanish colonists had become widespread before the 1850s, and much of the original oak woodland canopy was cut or burned off by the 1890s. For most California oak woodlands, the earliest aerial photography dates to the 1930s and very little ground-level photography was taken before the 1880s. Thus, the photographic record begins after major changes were completed. Earlier written accounts can add some information, but specific details on the floristic composition of oak woodlands are rare.
3. How have management practices and other human activities shaped current site conditions? Although information gaps will typically exist, historical site analysis can reveal important details about the succession of human impacts that have shaped the current landscape. Because site specific historical data is difficult to obtain, it is often necessary to rely on more regional historical information. However, the history of land ownership and management for many parcels is so complex that one should generally not assume that past management has been uniform across wide areas.
The analysis of past and current vegetation and management can help one determine what outcomes are possible for a site, and what inputs may be necessary to achieve these outcomes. A key question to be addressed in this analysis is whether a site can still support the type of woodlands that were historically present. For example, valley oak is found where its roots access relatively shallow water tables or exploit a large reservoir of available capillary water stored in the soil profile. In historic valley oak sites where precipitation and soil available water holding capacity are low, significant lowering or elimination of shallow water tables may render the site unsuitable for this species.
What do you want?
Goals for oak woodland restoration
The overall goal of ecological restoration is to return an ecosystem to a former condition that includes the entire complement of plants and animals and the dynamic processes found in the naturally-occurring state. Presettlement conditions are the benchmark for the natural state, but we cannot define these conditions with certainty. While we remain unsure about the natural condition of the oak woodlands and the processes that maintained them, we can be fairly certain that restoring these woodlands to the presettlement state is virtually impossible. Thus, the goals we set for oak woodland restoration and management must be more modest than the goal of complete ecological restoration. The following goals are attainable because they acknowledge limitations due to the current degree of ecological degradation and our lack of knowledge about the pristine ecosystem.
1. Reestablish appropriate oak species in areas that did or could have supported oak woodlands previously and are now capable of supporting this vegetation type.
2. Establish sustainable populations of historically known and likely indigenous plant species and associations within oak woodlands.
3. Manage remnant oak woodlands and restored stands to permit natural regeneration and maximize the cover and dominance of indigenous plant species while minimizing the cover of nonindigenous species.
4. Promote reestablishment of natural biotic systems, including interacting microbial, invertebrate, and vertebrate communities, within restored woodlands.
Ultimately, all four goals should be met in a successful restoration. However, it may not be possible to effectively address all goals in the initial phase of a restoration project. Because oaks provide structure and canopy influence that drives both vegetation and wildlife dynamics in oak woodland ecosystems, establishing oak canopy is usually the initial goal addressed in a restoration project.
Prioritizing restoration sites
Although the need for oak woodland restoration is great, financial resources available for undertaking oak woodland restoration will always be limited. To make the best use of limited resources, it is desirable to compare the costs and benefits of potential project locations and technical approaches. Such an analysis would ideally be done on a regional basis, but issues of land ownership, land use, and sources of funds immediately pose constraints that restrict site consideration. Consequently, prioritization is typically conducted among lands under a common ownership (e.g., by a public park district) and on individual parcels.
Potential restoration sites can be prioritized according to existing oak resources, site quality, project benefits, and long-term sustainability. These criteria take into account the relationship of the restored area to the landscape in which it exists, and how it will influence or be influenced by existing oak woodlands in the area.
1. Presence of oaks on the site and proximity to existing woodlands. Existing woodlands and oaks in and near restoration sites are a likely source of locally-adapted plant germplasm as well as a potential reservoir of other organisms endemic to oak woodlands. Seeds and other propagules of understory plants from adjacent oak woodlands can be transported to restoration sites through the action of animal vectors, wind, and water. The passive movement of native nonvascular plants, lichens, fungi, actinomycetes, and bacteria into the restoration site is also favored if oak woodlands adjoin the restoration site.
2. Site quality. Site quality is the ability of a site to support oak establishment and growth. Site quality is primarily related to physical factors such as soils, climate, hydrology, and topographic position. However, oak survival and growth are also constrained by competing vegetation, herbivores, and disturbance factors that affect overall site quality for restoration purposes. Faster establishment of oak woodlands can be achieved by focusing initial restoration efforts on high quality sites. Furthermore, restoration costs are normally lower per unit area on high quality sites than on low quality sites.
3. Site-specific benefits. Ecological, societal, and economic benefits provided by oak woodland restoration can vary greatly between sites. Depending on soil type and topography, benefits such as erosion protection and soil stabilization may be significant or not. Restoration projects near migration corridors or critical habitat for species of special concern may provide greater benefits for wildlife habitat. Restored woodlands near urban areas may provide more direct benefits to human populations (recreation, modification of urban climates, hydrologic effects) than woodlands in remote areas.
4. Likelihood of long-term sustainability. Restored woodlands that will require few or no additional inputs after the original project will typically provide maximum benefits for minimum cost. Overall, prospects for sustainability result from a combination of site qualities (e.g., existing populations of understory plants), management (e.g., grazing practices), and land ownership and use that confer a degree of long-term stability (e.g. natural reserves or park lands). Reserves owned by public agencies or private land conservation organizations may ensure long-term protection against changes in land use, but projects on these lands may not be self-sustaining over the long term if management practices are poor or site quality is marginal.
How do you get what you want?
Factors that constrain regeneration
Oak woodland restoration is necessary where natural regeneration has failed in the past or is currently failing. The first task in planning a restoration project is a site analysis to determine what factors are constraining natural regeneration of oaks and associated plants. Suitable sites for restoration are those in which site management has inhibited oak woodland regeneration but edaphic and climate factors are not critically limiting for seedling establishment.
The influence of management on oak reproduction is illustrated by the fact that oak seedlings volunteer readily in horticultural landscapes and along roadsides beyond pasture fences. These environments constitute safe sites for oak reproduction. Many of the numerous differences that exist between adjacent garden and rangeland environments are directly related to the ease with which oak seedlings establish in the former environment but not the latter. Differences in acorn dispersal, seedbed conditions, herbivory, soil moisture, shading, and fire frequency influence seedling establishment. As discussed below, one or more of these factors may critically constrain natural regeneration at a restoration site.
Acorn dispersal
Most acorns that fall land under or near maternal tree canopy. If seedbed conditions are favorable, some of the acorns that are not eaten or carried off by animals will germinate and become part of a persistent seedling bank. In blue oak (Swiecki and Bernhardt 1998) and probably other California oaks, this seeding advance regeneration is suppressed by overstory trees. Although such seedlings can persist many years in the understory, they are not recruited to sapling or tree size classes unless overstory competition is reduced though decline, mortality, or removal of the oak overstory. Hence, gap-phase replacement is a primary mode of reproduction in at least some California oaks.
Establishment of oak seedlings well beyond the maternal canopy depends primarily on dispersal by animals. The California scrub jay (Aphelocoma californica) is probably the most important acorn vector due to its abundance, wide distribution, and ability to cache thousands of acorns in a season. Because scrub jays typically bury their acorns in sites with loose soil and/or a layer of organic debris, unrecovered acorns are well-positioned to germinate and develop into seedlings. Scrub jays do not randomly place acorns in grasslands, but typically cache acorns near landmarks such as fencelines, rock outcrops, trees, and shrubs. Hence, reinvasion of grasslands by oaks can be limited in part by a lack of acorns.
In riparian systems, floodwaters may also be an important means of long-distance dispersal. Dam construction has eliminated annual flooding events from most California rivers, largely eliminating flood flows as a major acorn dispersal method.
Seedbed conditions
Most California oaks do not require stratification and begin to germinate as soon as moisture is available in the autumn. However, acorns may fall several months before the onset of the winter wet season. Unless acorns are buried or protected by leaf litter, they desiccate and lose viability during the warm dry conditions typical of late summer and autumn. In nongrazed woodlands, a substantial layer of organic debris accumulates on the soil surface beneath oak canopies. Acorns falling from the canopy readily penetrate into this duff layer where they are largely protected from desiccation, overheating, and to some degree, from vertebrates. Because long-term grazing reduces or eliminates the litter layer beneath trees and compacts soils, most acorns in grazed lands remain exposed on the soil surface where they desiccate or are consumed by livestock and other vertebrates. Sudworth (1908) noted that poor seedbed conditions inhibited reproduction of several oak species in both grazed and agricultural lands.
Herbivory
Herbivory can severely limit the growth and survival of oak seedlings and saplings. Livestock, deer, and rodents all have the potential to limit or eliminate oak reproduction, but the relative importance of each herbivore varies by location. Livestock, most commonly cattle, are the most important herbivore limiting oak regeneration over the greatest proportion of California’s oak woodlands. Browsing and trampling by cattle shortens the life of individual seedlings and can deplete or eliminate understory advance regeneration. Cattle browsing can also indefinitely suppress the growth of seedlings located beyond the canopy that would otherwise recruit to sapling and tree size classes. By prolonging the period that juvenile oaks remain in small size classes, herbivory increases the susceptibility of oak regeneration to both subsequent herbivory and fire. The impact of livestock herbivory varies somewhat by species. The less palatable evergreen oaks Q. agrifolia and Q. wislizeni are less severely browsed than valley oak and blue oak.
Gophers, ground squirrels, and voles can kill juvenile oaks by chewing and girdling stems. Elimination of predators and alterations in vegetative composition and structure can increase rodent populations to the point that they can inhibit oak reproduction. Rodent populations vary spatially and can fluctuate from year to year due to habitat conditions, predators, and pathogens. Because rodents are not problems at all locations, observations at the restoration site are needed to determine whether rodent herbivory is likely to be a significant constraint. Insects, particularly grasshoppers (Melanoplus spp.) sometimes cause significant damage to young oak seedlings, but insect herbivory is not usually a severe constraint to oak reproduction (Swiecki et al. 1991).
Soil moisture
Water stress associated with summer drought also limits oak seedling survival and growth. Water stress effects are most acute at the early seedling stage. As oak seedlings become established and develop an extensive root system, water stress is less likely to cause mortality, although it may limit growth rates and thereby prolong the period during which seedlings are susceptible to herbivores or fire. Seedling tolerance to water stress varies between oak species.
Vegetation influences the level of water stress to which oak seedlings are exposed in several ways. Overstory and understory plants compete with oak seedlings for available soil moisture. Overstory and tall understory species can also shade oak seedlings, elevate relative humidity, and reduce temperature and wind speed, thereby reducing evapotranspiration demand. Many California plant ecologists accept the conjecture that soil moisture is less available in oak woodlands now than it was in presettlement times due to the replacement of native herbaceous vegetation with nonindigenous annual grasses and forbs. Definitive evidence in support of this hypothesis is lacking, largely because the nature of the presettlement understory is poorly understood. Blue oak seedlings experience high levels of water stress during the summer (Griffin 1973), but it is not possible to determine whether these levels of summer water stress have changed since settlement.
Insolation
In open woodlands and clearings, high levels of insolation (solar radiation) function mainly to increase evaporative demand and consequently, water stress. In relatively xeric sites and/or dry years, high levels of insolation can limit seedling survival in some oak species (Muick 1997, Borchert et al. 1989). In some xeric areas, the density of existing second-growth oak stands is greater on northerly aspects or is entirely restricted to north slopes, indicating that insolation has strongly affected regeneration since settlement.
Oak seedling and sapling growth and survival can also be limited by a lack of light in woodlands with closed or nearly closed canopies. Levels of shading that inhibit oak growth and survival vary by growth stage, site conditions, and oak species. Blue oak seedlings can establish under canopy, recruitment to the sapling stage typically requires a canopy opening (Swiecki et al. 1997b). Blue oak saplings that become overtopped by faster-growing species usually decline and die. Although shady conditions generally favor seedling establishment in xeric sites, shade may reduce seedling establishment if soil moisture is not limiting (Borchert et al. 1989).
Fire
Most California oaks possess one or more adaptations that allow them to tolerate infrequent fires. Mature trees of some species, including Engelmann, blue, and valley oak, tolerate light to moderate ground fires with little damage. However, more intense fires may kill trees outright or create fire scars that facilitate invasion by wood decay fungi and lead to early mortality. Virtually all young California oaks resprout readily after topkill by fire, and some species, including Q. agrifolia and Q. wislizeni, crown sprout vigorously after topkill even as mature trees.
In mesic areas where black oak (Q. kelloggii) or Oregon oak (Q. garryana) coexist with fire-sensitive conifers, infrequent fires may be important in suppressing succession to coniferous forest and maintaining oak as the dominant canopy species. However, these situations are the exception rather than the rule in California oak woodlands. In most low elevation woodlands, fire is not required for regeneration or to maintain the dominance of the oak overstory.
Even though oak seedlings and saplings resprout readily after topkill, fire causes low to moderate levels of mortality in juvenile oaks. After topkill, juveniles may require at least several years to recover their aboveground biomass (Figure 1). Repeated destruction of the shoot in successive years depletes seedling energy reserves thereby increasing the incidence of fire-related mortality and reducing sapling recruitment (Swiecki et al. 1997b). Frequent fire suppresses oak reproduction and facilitates conversion of woodlands and savannas to grasslands, a pattern seen in other areas. The combination of repeated fire and grazing is especially effective for suppressing regeneration, and was historically used to convert woodlands to grasslands.
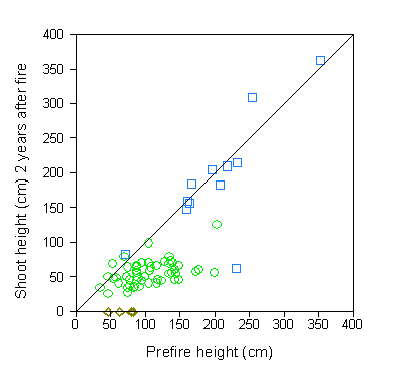
Figure 1. Effect of fire on survival and height growth of natural juvenile blue oaks in a grassland dominated by introduced annual grasses. Shoot heights on the Y-axis were recorded 2 years after the fire. Points below the diagonal line represents oaks that have not regrown to their pre-fire height.
Addressing constraints with restoration inputs
At a given site, one or more of the constraining factors listed above may be inhibiting seedling establishment and growth. The minimum restoration inputs needed to restore oaks at a given site are those that address the critical site constraints, i.e., factors that completely inhibit plant establishment. Most restoration inputs have multiple effects on the ecosystem and can change the intensity of several constraints, either positively or negatively. An integrated approach is needed to balance the positive and negative influences of selected inputs in the design of a restoration project.
Each restoration input also has corresponding costs in terms of labor and materials, as well as possible ancillary costs (e.g., disposal of used materials). Although some inputs may provide statistically significant differences in survival and growth, they may not necessarily be cost effective. To achieve a high level of cost effectiveness, not only should inputs be matched to site constraints, but the least expensive effective inputs should be selected.
Planting
Genetic considerations
Locally-collected seed is recommended for restoration plantings for two interrelated reasons. First, local genotypes are likely to be well-adapted to local soil and climate conditions, and therefore are likely to perform well. The importance of using local ecotypes to ensure seedling survival and growth is well documented for commercial conifer species, and evidence for local adaptation has been demonstrated for northern red oak (Quercus rubra) from the eastern USA (Sork et al. 1993). Secondly, plantings from non-local seed may contaminate the local gene pool by introducing maladaptive alleles into the local population. Through outcrossing with non-local individuals, specialized traits of local populations may be compromised or diluted.
Unfortunately, little is known about the genetics of California oak species. Phenotypic variability between oak populations has long been noted in California (Jepson 1910). The common occurrence of interspecific oak hybrids indicates that gene flow between species may contribute substantially to variation between local oak populations (Dodd et al. 1997). Several studies have examined the amount of phenotypic variation that exists within and between populations of in several oak species (e.g., Rice et al. 1993,1997). However, current data do not indicate how large a local seed collection zone should be, or whether oaks growing on hilly terrain differ genetically from oaks of the same species growing on adjacent valley floors.
For at least some California oak populations, we may not be able to assume that local germplasm is highly adapted to current site conditions. If site conditions are more xeric now than they were prior to settlement, oak genotypes from a more xeric location (e.g., lower elevation and/or latitude) might be better adapted to the site than the extant local population. For example, initial results from a reciprocal transplant test indicate that blue oak seedlings from a distant xeric site (Sierra Nevada foothills) performed better than the local germplasm at a relatively mesic site (North Coast range) (Rice et al. 1997).
Defining local seed is further complicated by the movement of oak germplasm between distant populations by humans. Along aboriginal routes of migration and trade, human-assisted gene flow may have significantly affected population genetics of oaks and other species used for food. Recently, the increased horticultural use of California native oaks in urban and rural landscaping, including highway plantings, has distributed many non-local oaks amid oak woodlands. In some areas, acorns collected from local trees may not represent germplasm of a locally-adapted population.
Until further genetic studies are completed, the designation of the local seed area for each species must be guided by a knowledge of the restoration site and local woodlands. In practice, woodlands within the same watershed located within a few to perhaps 10 km of the restoration site may be considered likely candidates for local germplasm. Microclimate and soils of the source site should match the target site to the degree possible. Collections from several source areas representing as many trees as possible should be used to obtain greater levels of diversity in the germplasm. Acorns should not be collected from trees likely to be pollinated by horticultural oaks of unknown provenance. In areas where oak populations have been extirpated or reduced to a few remnant individuals, truly local germplasm may not exist or may have insufficient levels of genetic diversity. In such situations, introduction of seed from more distant areas may be necessary.
Planting stock: seed vs. transplants
Direct planting of acorns has several significant advantages over transplanting even though oak seedlings can be successfully established by either method. Unlike acorns, transplants require space for propagation and care in the nursery. Transplants are more difficult to store and transport, and require more effort and care in planting than acorns. When direct-seeded in the field, seedlings of many California oak species produce a long taproot which can extract moisture from deep in the soil profile (Matsuda et al. 1989). In both container-grown and bareroot nursery stock, the dominance of the taproot is destroyed, impairing the drought tolerance of the developing seedling. Consequently, transplants normally require some irrigation, whereas direct-seeded acorns do not. Finally, nonindigenous soil-borne pathogens or insect pests from the nursery may be introduced with the transplants into the planting site; this risk is negligible for direct-seeded acorns. For most restoration plantings, transplants have no long-term survival or growth advantages over direct seeded acorns that would offset their increased cost and other drawbacks.
The choice of planting stock also has genetic implications. Direct-seeded acorns are subjected to site-specific selection pressure at the earliest possible stage of growth. This facilitates selection for seedling characteristics that may contribute to fitness, including resistance to insects and other herbivores, drought and temperature tolerance, emergence date, the structure and growth rates of roots and shoots, and response to local mycorrhizal fungi and other rhizosphere microorganisms. Selection pressures imposed on oak seedlings grown as nursery stock are vastly different from those encountered at the restoration site and typically bear little relationship to field conditions. Hence, genotypes that are successful in the nursery may not be the same as those that would have succeeded in the field. Paradoxically, the main advantage of transplants, i.e., ensuring that every planting site contains a seedling, is a disadvantage in terms of allowing natural selection to function.
Planting practices
In addition to addressing inadequate acorn dispersal, planting ameliorates poor seedbed conditions. A suitable seedbed can be prepared by turning over and breaking up the upper 25-30 cm of the soil profile with a shovel. Augering and backfilling deep (60-90 cm) planting holes is more expensive and does not always provide a clear benefit (e.g., Figure 2), but may be beneficial where root penetration is inhibited by subsurface strata such as tillage pans or clay lenses. Deep augered holes can also settle excessively after wetting, which can be detrimental to seedling establishment.
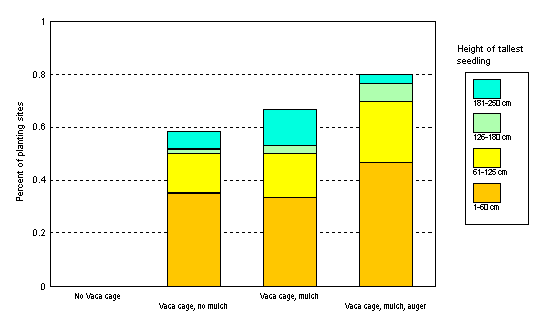
Figure 2. Percent of valley oaks in each of 4 height classes. Oaks were planted from acorns in 1989 (Bernhardt and Swiecki 1997, west hillside). The site (3 ha) is grazed annually at varying stocking levels. The season and duration of grazing has also changed from year to year, but usually is winter-spring. Nonwoven polypropylene landscape fabric (90 cm square) covered with waste wood chips was used at mulched sites. Ten years after planting, more than half of the surviving seedlings are still less than 60 cm tall. Vaca cages to protect seedlings from cattle grazing are essential for seedling survival at this site.
Oaks are strongly mycorrhizal, although the mycorrhizal fungi associated with California oaks are poorly characterized. Soil taken from existing woodlands can serve as a source of inoculum for mycorrhizal fungi and other beneficial soil microorganisms and invertebrates. Planting acorns with as little as 250 ml of woodland soil has been shown to have beneficial effects on seedling growth (Scott and Pratini 1997). Inoculating planting sites with woodland soil might be cost effective on some sites, but may not be necessary if potential sources of inoculum are close enough to permit natural colonization.
Fertilizer is considered to be an inexpensive input, but may be unnecessary or even counterproductive. In oak woodlands, soil moisture is typically more limiting than levels of mineral nutrients in the soil (Dahlgren et al. 1997). High levels of phosphorus can suppress mycorrhizal development, and excessive levels of nitrogen can increase susceptibility of plants to insect damage.
An organic mulch applied over the planting site moderates soil temperatures, reduces evaporative water loss, helps maintain high humidity around the seed, facilitates seedling emergence by preventing crusting of the soil surface, suppresses the growth of competing vegetation, and releases plant nutrients as it decays. Mulch containing chipped woody material may also provide a food base for woodland microorganisms that utilize woody debris. Such woody substrates are normally lacking in annual grasslands. Mulches can significantly improve early seedling growth and survival (Bernhardt and Swiecki 1997), but it is unclear whether long-term survival and growth benefits are sufficient to make this input cost effective for all sites (Table 1).
Table 1. Survival and relative costs of different sets of restoration inputs for valley oaks planted from acorns in 1989 (Bernhardt and Swiecki 1997, west hillside) and shown in Figure 2.
|
Treatment |
Survival in 1999 |
Set up worker-hours per treatment |
Initial (1989) material cost |
Setup hours/ |
Initial material cost/surviving site |
|
No Vaca cage, no mulch |
0 |
.23 |
0 |
- |
- |
|
Vaca cage, no mulch |
57% |
0.62 |
$5.36 |
1.06 |
$9.19 |
|
Vaca cage, mulch |
67% |
1.01 |
$6.40 |
1.51 |
$9.6 |
|
Vaca cage, mulch, auger |
80% |
1.14 |
$7.32 |
1.43 |
$9.15 |
Some highly effective planting techniques entail little or no cost. Seedling emergence rates can be improved by inspecting acorns at planting and discarding those with evidence of insect damage or decay. Planting several acorns per site increases the probability of having at least one successful seedling per site (Swiecki and Bernhardt 1991). Planting early, immediately after the first autumn rains, ensures that germinating acorns can take full advantage of seasonal rainfall. Shoot growth in the first two seasons after planting is greatest for acorns planted at the earliest possible date (McCreary 1990). Planting at an adequate depth (about 5 cm) protects acorns from desiccation and reduces the likelihood that they will be eaten by rodents (Tietje et al. 1991). Seedling survival and growth rates can be also improved by selecting favorable planting sites. For example, damage by rodents such as ground squirrels can be reduced by simply avoiding areas with active colonies (Bernhardt and Swiecki 1997).
Soil moisture limitations can be minimized by avoiding drought-prone areas, such as slopes with southwestern aspects or shallow soils, and planting preferentially in areas with naturally high soil moisture. In one planting, we observed that growth differences associated with highly favorable soil conditions far exceeded the effects of mulching and augering (Bernhardt and Swiecki 1997). It may not be possible to identify all of the most favorable microsites or restrict planting to them, but one can make better use of limited resources by avoiding obvious problem areas.
At any site, a given set of inputs may give rise to a range of outcomes depending on the year. In projects that require oak planting, a useful hedge against temporal stochastic constraints (e.g., extended drought, accidental fire, herbivore population fluctuations) is to plant in a number of successive years. This tactic has the effect of averaging risks over time and increases the chance that favorable growing conditions will coincide with at least some plantings.
Protection from herbivores
Livestock
Whether oak woodland restoration relies on natural regeneration or planting, it cannot succeed in areas that are heavily grazed by livestock (Figure 2). Long-term livestock grazing is highly destructive to oak woodland ecosystems because its adverse effects extend far beyond herbivory and trampling of oak seedlings. Livestock deplete the acorn supply and degrade seedbed conditions by removing litter and compacting soil. Excessive livestock grazing also increases populations of many nonindigenous annuals. Some of these (e.g., yellow starthistle, Centaurea solstitialis) extract water late in the season from deep in the soil profile, increasing competition for soil moisture. Potentially negative effects on the soil microflora and invertebrate fauna from compaction and changes in the herbaceous layer are also likely but are largely undocumented.
Cessation or strict limitation of grazing may be the only restoration input necessary where livestock grazing is the primary factor suppressing oak reproduction. Reduction in grazing intensity may be accomplished by reducing the season of use, animal stocking levels, and/or grazing frequency (e.g., rest rotations with no grazing in certain years). The maximum amount of grazing that is compatible with oak woodland restoration will vary greatly by site, but mesic sites will generally tolerate higher levels of grazing pressure than xeric sites.
Without continual suppression by livestock browsing, existing seedlings in open, noncanopied positions will recruit to the tree stage, but the length of time required will vary with site quality, oak species, and plant condition. Seedlings that have been suppressed for an extended period may require at least several years to reestablish a vertical leader. Removing or severely restricting grazing can also permit the recovery of understory seedling advance regeneration under oak canopy, other suppressed indigenous plant species, and soil ecosystem processes.
Many of the most aggressive nonindigenous weeds in California oak woodlands and savannas are favored by open sites, soil disturbance, and selective removal of competing vegetation by grazing animals. In time, many of these species will decline in dominance after grazing is eliminated and an oak overstory is reestablished. However, over a shorter time horizon, undesirable changes in the herbaceous layer may develop that may require management with tightly controlled grazing, herbicides, or other techniques. In northern California, the introduced perennial Harding grass (Phalaris aquatica) is suppressed in open sites by grazing but can develop into dense, highly competitive stands if grazing is removed (Bernhardt and Swiecki 1997). The short-term response of understory vegetation to a release of grazing pressure is largely site specific, and depends on existing populations of native and nonindigenous plants, site conditions, weather patterns, and management practices.
Reduced grazing pressure can also influence wildlife populations in ways that may affect the restoration process. Ground squirrel populations tend to be higher in grazed than in ungrazed locations, so reduced grazing may reduce damage caused by this species. Where cessation of grazing leads to heavy grass cover, vole (Microtus californicus) populations may increase to levels that reduce oak seedling establishment. Although the long-term prospects for restoring the oak woodland ecosystem are vastly improved when grazing is reduced or eliminated, close monitoring and active management may be required to minimize negative changes during the transitional period.
In areas where livestock grazing cannot be eliminated or adequately restricted, it is still possible to protect individual oak seedlings or saplings from browsing by using single tree exclosures. Individual exclosures must be relatively sturdy to withstand the abuse of cattle that pull at protruding oak branches and rub against the exclosures. For the past 10 years, we have successfully used a low-cost single-plant exclosure of our own design (Vaca cage) to protect both existing oak saplings and new planting sites from cattle (Swiecki and Bernhardt 1997, Figure 3). Vaca cages have been effective in protecting individual oaks or planting sites from cattle (Table 1, Figure 2). Periodic inspection, repair, and height adjustment is necessary to maintain the cages’ effectiveness. Cages must eventually be removed to prevent girdling and scarring of oak branches by cage wires.

Figure 3. Vaca cages used to protect direct-seeded valley oaks from seasonal cattle grazing. Even though oak growth is relatively rapid at this favorable site and cattle are only present for several months each year, below browse line nearly all branches outside of the cages have been destroyed. Cage heights can be extended to help saplings grow above browse line.
Selective protection of individual planting sites is a useful interim restoration tactic that does not require changes in the existing grazing regime. However, Vaca cages are relatively expensive and time-consuming to use, and do not protect other desirable understory vegetation or other elements of the ecosystem. If a site is excessively grazed, the ecosystem will remain highly degraded even if some oak trees are established though the use of Vaca cages.
Other herbivores
As noted above, management of herbaceous vegetation can significantly affect rodent populations and damage at a restoration site. In localized areas, direct reduction of rodent populations by trapping, baiting, or other means may be practical. Alternatively, rodents can be excluded from the individual planting sites through the use of wire-mesh or aluminum screen cylinders or plastic tree shelters (McCreary and Tecklin 1997). Although the these protective devices are relatively inexpensive, substantial amounts of labor are required to install, inspect, and eventually remove the devices. Cost-effectiveness is therefore likely to be low unless rodent populations are critically limiting.
Deer browsing can stunt oak seedlings and saplings, but damage caused by deer is typically less severe than that caused by cattle (Bernhardt and Swiecki 1997). We have successfully used inexpensive cylindrical cages made of galvanized poultry netting to protect seedlings from deer. Such cages usually do not require maintenance other than eventual removal, but would only be cost effective where deer browsing pressure is intense. Deer are classified as game animals, and management of local populations by hunting may be sufficient to reduce their impact to a tolerable level in some cases.
Minimizing moisture stress
Vegetation management
Moisture stress can be reduced by managing competing vegetation in the immediate vicinity of the oak seedling. Maintaining bare soil in a zone at least 60 cm in diameter for one or two seasons increased survival and growth of blue oak seedlings in the first 5 years after planting (McCreary and Tecklin 1997). Bare soil areas may be produced by mechanically scraping all vegetation off the soil surface (scalping), although this method is labor intensive because repeated treatments are needed. One or two properly timed applications of a nonselective foliar herbicide (e.g., glyphosate) can also be used to eliminate competing annual vegetation around an oak seedling (Tecklin et al. 1997), but the seedling must be shielded from the spray. Some soil-applied broadleaf herbicides can have negative effects on oak root growth and soil microorganisms, and should be not be used without thorough testing. Herbicide use and scalping typically result in bare, unshaded soil around the seedling which is subject to erosion and can impede percolation of rainfall into the soil. In contrast, mulch suppresses competing vegetation and reduces rainfall runoff and evaporation without increasing erosion potential.
Depending on the weedy species present at a site, it may be more efficient to manage herbaceous vegetation across the entire restoration site. Possible management choices include properly timed mowing, tillage, or herbicide applications. Such techniques are more likely to be feasible on sites with few existing desirable plant species and where negative environmental consequences are unlikely. Regular spacing of oaks in rows or grids has been used to facilitate mowing operations in some projects. Precisely-managed, limited duration grazing can be used to differentially remove palatable herbaceous species, but unless individual oak seedlings are protected, it is difficult to manage grazing so that the reduction in herbaceous competition outweighs herbivory. Grazing around protected seedlings can lead to higher growth and survival compared to adjacent nongrazed sites (Bernhardt and Swiecki 1997).
The practical use of fire for vegetation management in oak woodlands is restricted to situations in which competing vegetation is slower-growing and/or significantly more fire-sensitive than oak seedlings and saplings. Infrequent fires (e.g., 10-20 year intervals) may be useful in suppressing the encroachment of fire-sensitive conifers into certain mesic oak woodlands (Fritzke 1997, Hastings et al. 1997). However, the frequent fires needed to manage undesirable annuals would destroy oak regeneration and inhibit oak woodland restoration. In areas that burn frequently, oak regeneration can be favored by reducing fire frequency.
Different vegetation management approaches can vary widely in cost-effectiveness. Unless competing vegetation critically limits oak reproduction, it may be more cost efficient to forego vegetation management and accept low growth rates. Such an approach is more feasible where risks associated with herbivory and fire frequency are low.
Microclimate modifying devices
Plastic tree shelters not only protect oak seedlings from rodent herbivory, but also provide a seedling microclimate characterized by reduced light intensity and air flow and increased temperature, humidity, and carbon dioxide levels (Burger et al. 1992, Minter et al 1992). The tree shelter microenvironment has profound effects on seedling morphology, especially when relatively tall (>60 cm) tree shelters are used. Stem height growth is typically enhanced in tree shelters, but shoot growth responses vary between and within species (Plumb and DeLasaux 1997, McCreary and Tecklin 1997, Burger et al. 1997). Tree shelters reduce total plant biomass and the root:shoot ratio in at least some oak species (Burger et al. 1997), which could compromise long-term survival in some situations. Tree shelters have improved initial oak seedling survival in some, but not all studies (McCreary and Tecklin 1997, Plumb and DeLasaux 1997). Both shade and protection against rodent herbivory can contribute to seedling survival, so it is difficult to determine which effect predominates when tree shelters are used. Long-term results from restoration plantings are needed to accurately evaluate the costs and benefits of tree shelters.
Aluminum screen cages also provide a modified microenvironment, though less extreme than that within plastic tree shelters. Screen cages reduce incident sunlight by about half (Adams et al. 1991) and may slightly increase relative humidity in the immediate vicinity of the seedling by reducing wind velocity. Depending on their construction, screen cages can also provide protection against rodent herbivory, at least while seedlings are small. McCreary and Tecklin (1997) found that screen cages provided less protection against rodents and less of an enhancement of shoot growth than tree shelters, whereas Costello et al. (1996) found tree shelters and screen cages to be equally effective in enhancing shoot growth and survival.
Irrigation
Although irrigation can enhance oak seeding growth, it is a relatively expensive and high-maintenance input,, especially in plantings without nearby water sources. Hand watering is labor intensive and may be inefficient due to losses from evaporation and runoff. Drip irrigation systems allow better percolation of water into the soil profile but have higher equipment costs, especially if the water source requires extensive filtration. Labor required to install and maintain a drip irrigation system can also be high. We have observed sites where oaks became highly water-stressed when irrigation was discontinued, presumably due to effects of irrigation on root distribution and/or root:shoot ratios. Irrigated planting sites are also more likely to be attacked by ground squirrels and gophers than nonirrigated sites. Consequently, summer irrigation does not always improve oak seedling growth and survival and even early benefits of irrigation can be short lived (Swiecki and Bernhardt 1997). Hence, irrigation can be one of the least cost-effective inputs in a restoration project.
Protecting existing oak reproduction
Although most oak woodland restoration projects involve planting, planting is not always necessary to restore oak canopy. In high-quality sites adjacent to oak stands, suppressed natural seedlings and saplings may be present in the area targeted for restoration. If suppression of juvenile oaks is not due to competition with the tree canopy but is associated with livestock browsing, mowing, or frequent fire, shoot protection can allow these oaks to recruit to the overstory. Suppressed juvenile oaks typically have well-established root systems, and can exhibit high shoot growth rates once they are protected. Oaks may be protected individually (e.g., with Vaca cages) or across the entire site (e.g., by eliminating grazing). Growth rates of protected juvenile oaks vary by species and with site conditions, but a minimum of 5 to 10 years of protection is typically needed to recruit juvenile oaks to a size class for which further protection is unnecessary. By protecting existing juvenile oaks, one can restore oak canopy with locally-adapted material in a shorter time than is possible by planting.
Other species
Many plant species other than oaks are important components of oak woodlands. Some other tree species, such as California buckeye (Aesculus californica), can be direct-seeded in a manner similar to oaks and can easily be included in the initial phases of restoration. Other species that are good candidates for early phases of restoration are those that do well in open sites but will not compete excessively with oak seedlings. Perennials that can be propagated by root, corm, or bulb divisions but reproduce sparingly by seed should also receive priority for reintroduction, especially in situations where local populations may be threatened with extinction. Transplanting divisions from local perennial populations can help reintroduce associated soil microorganisms and invertebrates into the restoration site. Because little is known about the genetics of other oak woodland canopy and understory species, a conservative approach toward utilizing local material is warranted.
Are you getting what you want?
Even with proper planning, factors that influence the success of restoration efforts can vary widely across space and time, and many of these factors are beyond the control of the restorationist. Outcomes for a given set of restoration inputs may vary by year, location, and species. Because our ability to predict project outcomes is limited, every restoration project is experimental. Replicated trials of specific techniques and appropriate monitoring of restoration projects over an extended time period are needed to determine whether inputs have been effective over the long term. If labor and materials costs are carefully tracked, it is also possible to determine which inputs are cost effective. This empirical data can be used to adjust inputs at the restoration site and can be used to help design future projects.
Slow oak growth rates are typical in many California oak woodlands and savannas. In nonirrigated valley oak restoration plantings protected from cattle browsing, we have observed average shoot height increases of 5 to 10 cm per year or less in upland sites of moderate quality over the first 5 to 10 years after planting. For a fast-growing species like valley oak on a favorable site, a sizeable canopy tree can develop in about 25 to 30 years. For blue oak, a slower-growing species that typically occurs in relatively xeric sites, 100 years may be required to produce a tree with a moderate canopy. With the exception of some riparian floodplains that have been removed from agricultural use, most land available for restoration is of relatively low site quality. Restoring oak canopy and understory vegetation, along with functioning natural regeneration processes, will clearly take a long time. Long-term monitoring is necessary to evaluate the success of oak woodland restoration, but few restoration projects are more than 10 years old, and few of these have been monitored beyond 3 to 5 years.
Many mandated oak restoration projects have a horticultural bias, and high initial survival rates are considered to be evidence of success. These projects often use relatively large nursery stock and maximize restoration inputs in an attempt to reduce all possible constraints to the point of insignificance. This tactic may not be cost-effective, because restoration can succeed without addressing constraints that only partially reduce growth or survival. Furthermore, the horticultural approach can eliminate natural selection for seedling characteristics that confer fitness. If seedlings vary genetically and we allow natural selection to function in the restoration process, high rates of seedling mortality can be expected in a successful restoration. If our primary restoration goal is to establish self-sustaining woodlands, it may be unwise to thwart selection for adaptive seedling characteristics through the excessive use of horticultural inputs.
Conclusions
Oak woodland restoration is in its infancy in California. Most restoration projects undertaken to date have been limited to the establishment of oak trees. Growing California oaks is not a technically difficult task in a controlled situation, but most areas in need of restoration are not subject to tight control. Furthermore, growing a few oaks can be a relatively easy task, but establishing thousands of oaks over large areas is an arduous undertaking. Restoration of the oak canopy can be considered successful once a naturally-regenerating stand is established, which may require a century or more. Although oaks are the dominant and most important element of oak woodlands, we cannot consider oak woodland ecosystems to be restored if other important vegetation and wildlife components are lacking. Oak woodland restoration may begin by planting oaks, but must eventually address the sustainable management of the entire ecosystem.
Current management practices, especially grazing, must be addressed throughout oak woodlands if large scale restoration of the ecosystem is to occur. By using a more holistic restoration prioritization and planning process, restoration efforts can be focused in areas where needs are high and costs can be minimized by working with natural processes. Further cost efficiency can be achieved by matching inputs to the site as closely as possible and avoiding unnecessary inputs. Restoration and management are not deterministic processes, but are influenced strongly by stochastic events and processes. Long-term monitoring of the ecosystem is needed to determine the outcome of restoration and management inputs and to provide the data needed to adjust inputs appropriately.
Literature Cited
Adams TE, Sands PB, Weitkamp WH, McDougal NK (1991) Blue and valley oak seedling establishment on California’s hardwood rangeland. In: Standiford RB (tech coord) Proceedings-Symposium on Oak Woodlands and Hardwood Rangeland Management. U.S.D.A. Forest Serv Gen Techn Rep PSW-GTR-126. Pacific Southwest Research Station, Berkeley, California; pp. 41-47
Bernhardt EA, Swiecki TJ (1997) Effects of cultural inputs on survival and growth of direct seeded and naturally occurring valley oak seedlings on hardwood rangeland. In: Pillsbury NH Verner J Tietje WD (tech coords) Proceedings-Oak Woodlands: Ecology, Management, and Urban Interface Issues. U.S.D.A. Forest Serv Gen Techn Rep PSW-GTR-160. Pacific Southwest Research Station, Albany, California; pp. 301-311
Block WM, Morrison MM, Verner J (1990) Wildlife and oak-woodland interdependency. Fremontia 18:72-76
Bolsinger, CL (1988) The hardwoods of California's timberlands, woodlands and savannas. Resour. Bull. PNW-148. Portland, OR: Pacific Northwest Research Station, U.S. D. A Forest Service 148 p
Borchert, MI, Davis FW, Michaelsen, J, Oyler LD (1989) Interactions of factors affecting seedling recruitment of blue oak (Quercus douglasii) in California. Ecology 70:389-404
Burger DW, Forister GW, Gross R (1997) Short and long-term effects of treeshelters on root and stem growth of ornamental trees. Journal of Arboriculture 23(2) 49-56
Burger DW, Svihra P, Harris R (1992) Treeshelter use in producing container-grown trees. HortScience 27:30-32
CalFlora Database (1998) UC Berkeley Digital Library Project. http://elib.cs.berkeley.edu:8080/calflora/ [Accessed June 1998]
Costello LR, Peters A, Giusti GA (1996) An evaluation of treeshelter effects on plant survival and growth in a Mediterranean climate. J Arboriculture 22:1-9
Dahlgren RA, Singer MJ, Huang X (1997) Oak tree and grazing impacts on soil properties and nutrients in a California oak woodland. Biogeochemistry 39:45-64
Dodd RS, Rafii SA, Kashani K (1997) Gene flow among populations of three California evergreen oaks. In: Pillsbury NH, Verner J, Tietje WD (tech coords) Proceedings-Symposium on Oak Woodlands: Ecology, Management, and Urban Interface Issues. U.S.D.A. Forest Serv Gen Techn Rep PSW-GTR-160. Pacific Southwest Research Station, Albany, California; pp. 127-133
Fritzke SL (1997) A California black oak restoration project in Yosemite Valley, Yosemite National Park, California. In: Pillsbury NH, Verner J, Tietje WD (tech coords) Proceedings-Symposium on Oak Woodlands: Ecology, Management, and Urban Interface Issues. U.S.D.A. Forest Serv Gen Techn Rep PSW-GTR-160. Pacific Southwest Research Station, Albany, California; pp. 281-287
Griffin JR (1973) Xylem sap tension in three woodland oaks of central California. Ecology 54:152-159
Hastings MS, Barnhart S, McBride JR (1997) Restoration management of northern oak woodlands. In: Pillsbury NH, Verner J, Tietje WD (tech coords) Proceedings-Symposium on Oak Woodlands: Ecology, Management, and Urban Interface Issues. U.S.D.A. Forest Serv Gen Techn Rep PSW-GTR-160. Pacific Southwest Research Station, Albany, California; pp 275-279
Jepson WL (1910) The silva of California. Memoirs Univ of Calif vol 2 Univ of Calif Press, Berkeley, California
Matsuda K, McBride JR, Kimura M. 1989. Seedling growth form of oaks. Annals of Botany 64:439-446McCreary DD (1990) Acorn sowing date affects field performance of blue and valley oaks. Tree Planters Notes 41(2):6-9
McCreary, DD Tecklin (1997) Effects of seedling protectors and weed control on blue oak growth and survival. In: Pillsbury NH, Verner J, Tietje WD (tech coords) Proceedings-Symposium on Oak Woodlands: Ecology, Management, and Urban Interface Issues. U.S.D.A. Forest Serv Gen Techn Rep PSW-GTR-160. Pacific Southwest Research Station, Albany, California; pp. 243-250
Mensing, S (1992) The impact of European settlement on blue oak (Quercus douglasii) regeneration and recruitment in the Tehachapi Mountains, California. Madrono 39:36-46.
Minter WF, Myers RK, Fischer BC (1992) Effects of tree shelters on northern red oak seedlings planted in harvested forested openings. North J Appl. For. 9:58-63.
Muick, PC (1997) Effects of shade and clipping on coast live and blue oak seedling mortality and growth in California annual grasslands. In: Pillsbury NH, Verner J, Tietje WD (tech coords) Proceedings-Symposium on Oak Woodlands: Ecology, Management, and Urban Interface Issues. U.S.D.A. Forest Serv Gen Techn Rep PSW-GTR-160. Pacific Southwest Research Station, Albany, California; pp. 135-145
Plumb TR, De Lasaux MD (1997) An evaluation of coast live oak regeneration techniques. In: Pillsbury NH, Verner J, Tietje WD (tech coords) Proceedings-Symposium on Oak Woodlands: Ecology, Management, and Urban Interface Issues. U.S.D.A. Forest Serv Gen Techn Rep PSW-GTR-160. Pacific Southwest Research Station, Albany, California; pp. 135-145
Rice JR Gordon DR Hardison JL Welker JM (1993) Phenotypic variation in seedlings of a keystone tree species (Quercus douglasii): the interactive effects of acorn source and competitive environment. Oecologia 96:537-547.
Rice KJ, Richards JH, Matzner SL (1997) Patterns and processes of adaptation in blue oak seedlings. In: Pillsbury NH, Verner J, Tietje WD (tech coords) Proceedings-Symposium on Oak Woodlands: Ecology, Management, and Urban Interface Issues. U.S.D.A. Forest Serv Gen Techn Rep PSW-GTR-160. Pacific Southwest Research Station, Albany, California; pp. 109-115.
Scott TA, Pratini NL (1997) The effects of native soils on Engelmann oak seedling growth. In: Pillsbury NH, Verner J, Tietje WD (tech coords) Proceedings-Symposium on Oak Woodlands: Ecology, Management, and Urban Interface Issues. U.S.D.A. Forest Serv Gen Techn Rep PSW-GTR-160. Pacific Southwest Research Station, Albany, California; pp. 657-660
Sork VL, Stowe KA, Hochwender C (1993) Evidence for local adaptiation in closely adjacent subpopulations of northern red oak (Quercus rubra L.) expressed as resitstance to leaf herbivores. Am. Naturalist 142:928-936
Swiecki TJ, Bernhardt EA (1991) Minimum input techniques for restoring valley oaks on hardwood rangeland. California Dept Forestry and Fire Protection, Forest and Rangeland Resource Assessment Program, Sacramento, California
Swiecki TJ, Bernhardt EA (1998) Understanding blue oak regeneration. Fremontia 26: 19-26
Swiecki TJ, Bernhardt, EA, Arnold RA (1991) Insect and disease impacts on blue oak acorns and seedlings. In: Standiford RB (tech coord) Proceedings-Symposium on Oak Woodlands and Hardwood Rangeland Management. U.S.D.A. Forest Serv Gen Techn Rep PSW-GTR-126. Pacific Southwest Research Station, Berkeley, California; pp. 149-155
Swiecki TJ, Bernhardt EA, Arnold RA (1997a) The California oak disease and arthropod (CODA) database. In: Pillsbury NH, Verner J, Tietje WD (tech coords) Proceedings-Symposium on Oak Woodlands: Ecology, Management, and Urban Interface Issues. U.S.D.A. Forest Serv Gen Techn Rep PSW-GTR-160. Pacific Southwest Research Station, Albany, California; pp. 543-552
Swiecki TJ, Bernhardt EA, Drake, C (1997b) Factors affecting blue oak sapling recruitment. In: Pillsbury NH, Verner J, Tietje WD (tech coords) Proceedings-Symposium on Oak Woodlands: Ecology, Management, and Urban Interface Issues. U.S.D.A. Forest Serv Gen Techn Rep PSW-GTR-160. Pacific Southwest Research Station, Albany, California; pp. 157-167
Swiecki TJ, Bernhardt EA, Drake C (1997c). Stand-level status of blue oak sapling recruitment and regeneration. In: Pillsbury NH, Verner J, Tietje WD (tech coords) Proceedings-Symposium on Oak Woodlands: Ecology, Management, and Urban Interface Issues. U.S.D.A. Forest Serv Gen Techn Rep PSW-GTR-160. Pacific Southwest Research Station, Albany, California; pp. 147-156
Tecklin J, Connor JM, McCreary DD (1997) Rehabilitation of a blue oak restoration project. In: Pillsbury NH, Verner J, Tietje WD (tech coords) Proceedings-Symposium on Oak Woodlands: Ecology, Management, and Urban Interface Issues. U.S.D.A. Forest Serv Gen Techn Rep PSW-GTR-160. Pacific Southwest Research Station, Albany, California; pp. 267-273.
Tietje WD, Nieves SN, Honig JA, and Weitkamp WH (1991) Effect of acorn planting depth on depredation, emergence, and survival of valley and blue oak In: Standiford RB (tech coord) Proceedings-Symposium on Oak Woodlands and Hardwood Rangeland Management. U.S.D.A. Forest Serv Gen Techn Rep PSW-GTR-126. Pacific Southwest Research Station, Berkeley, California; pp. 14-20.